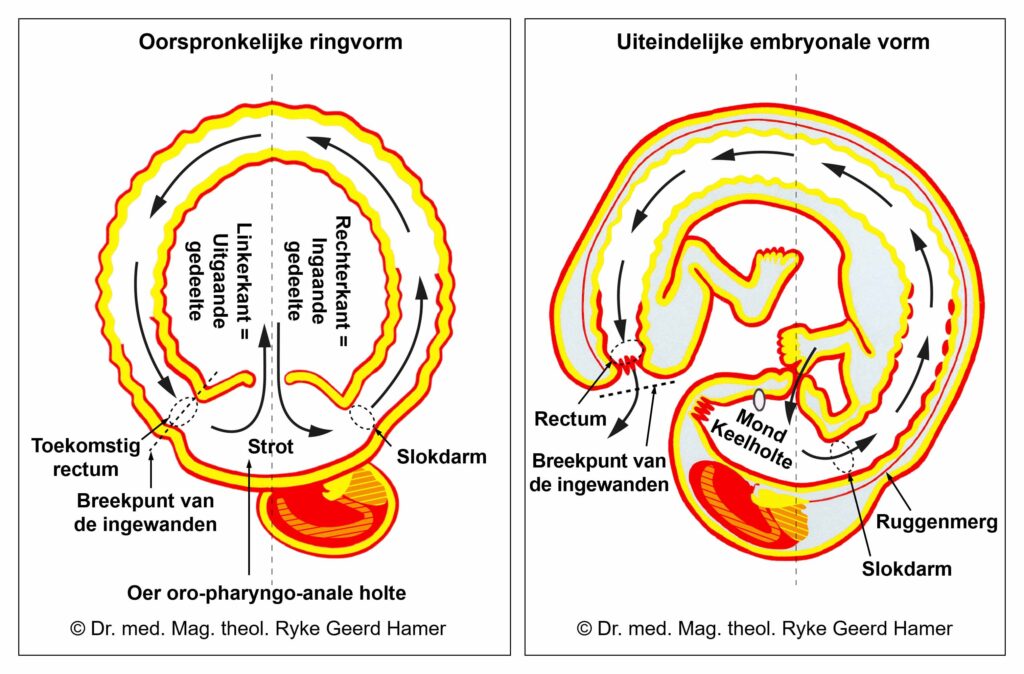
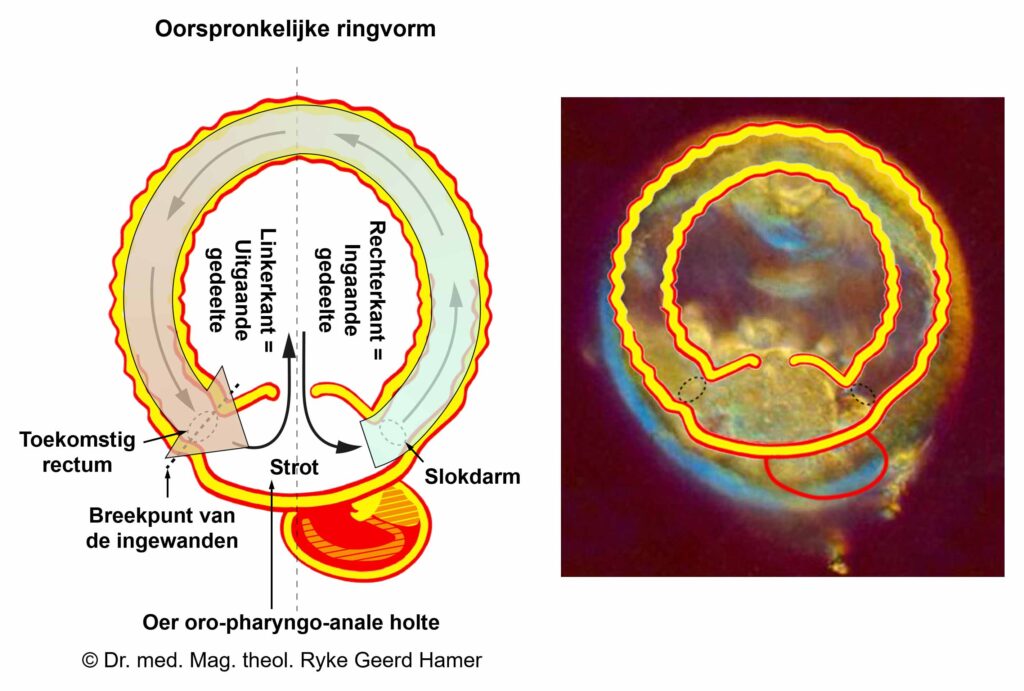
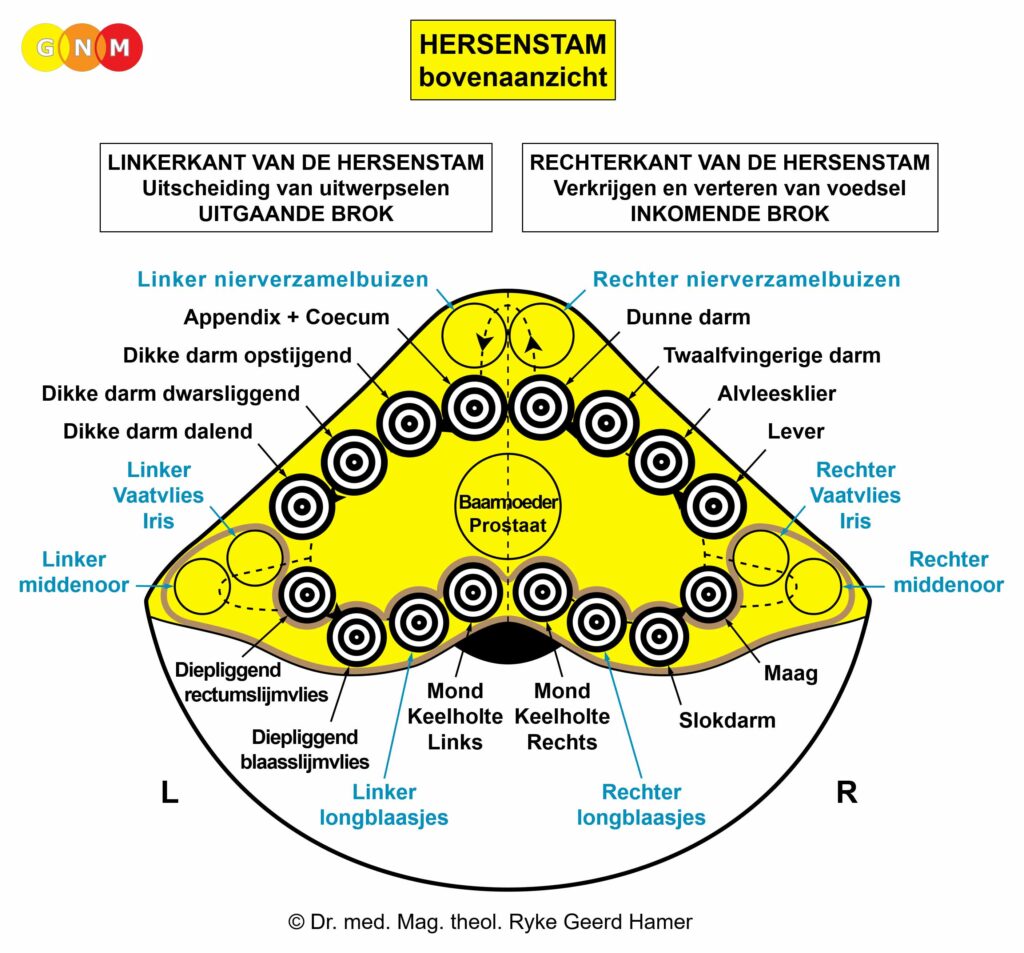
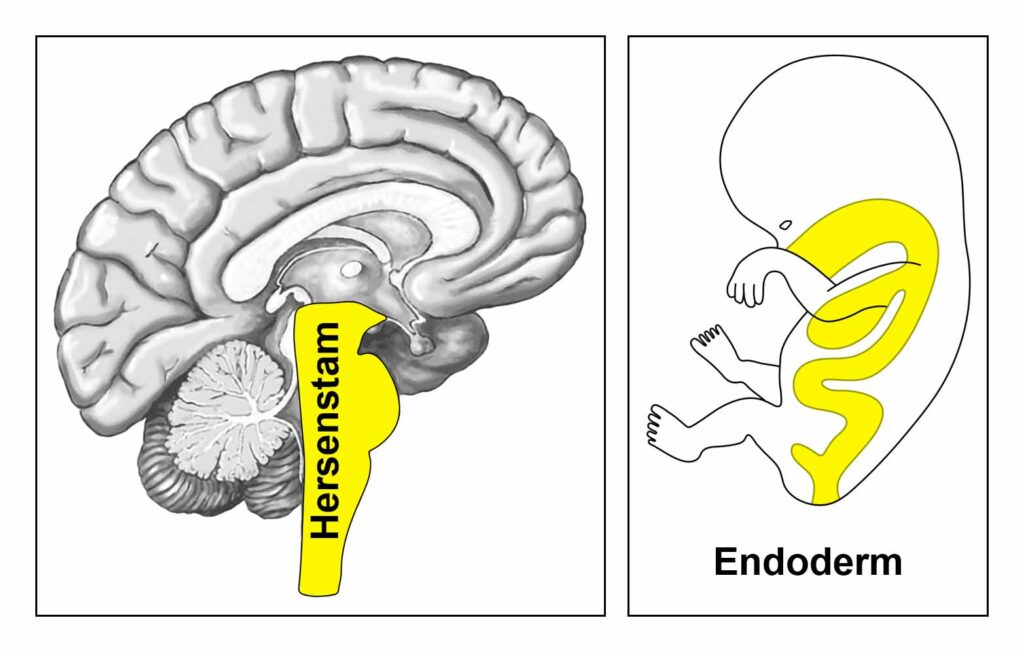
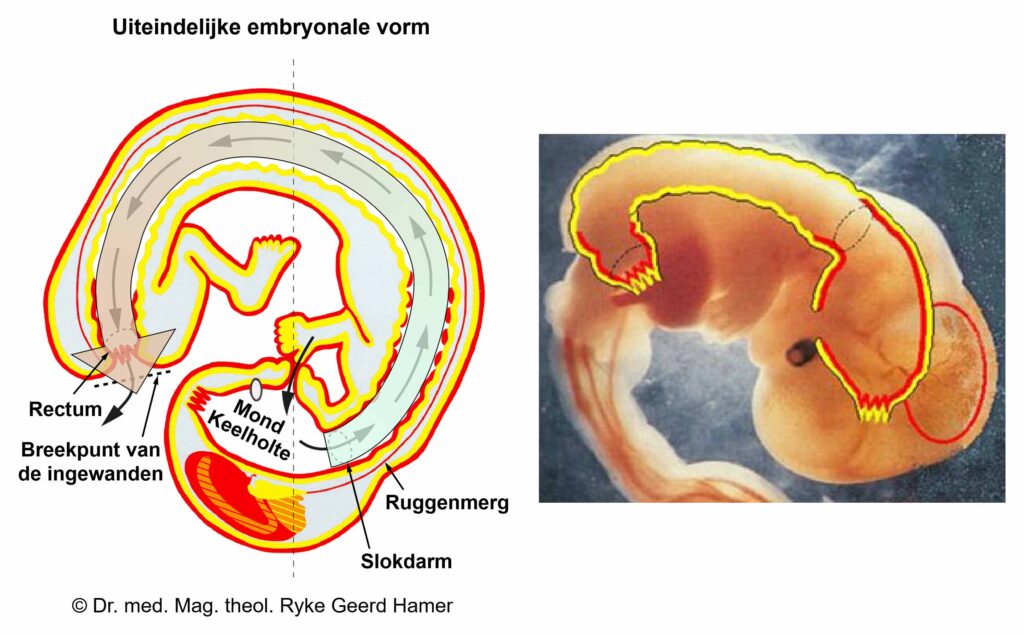
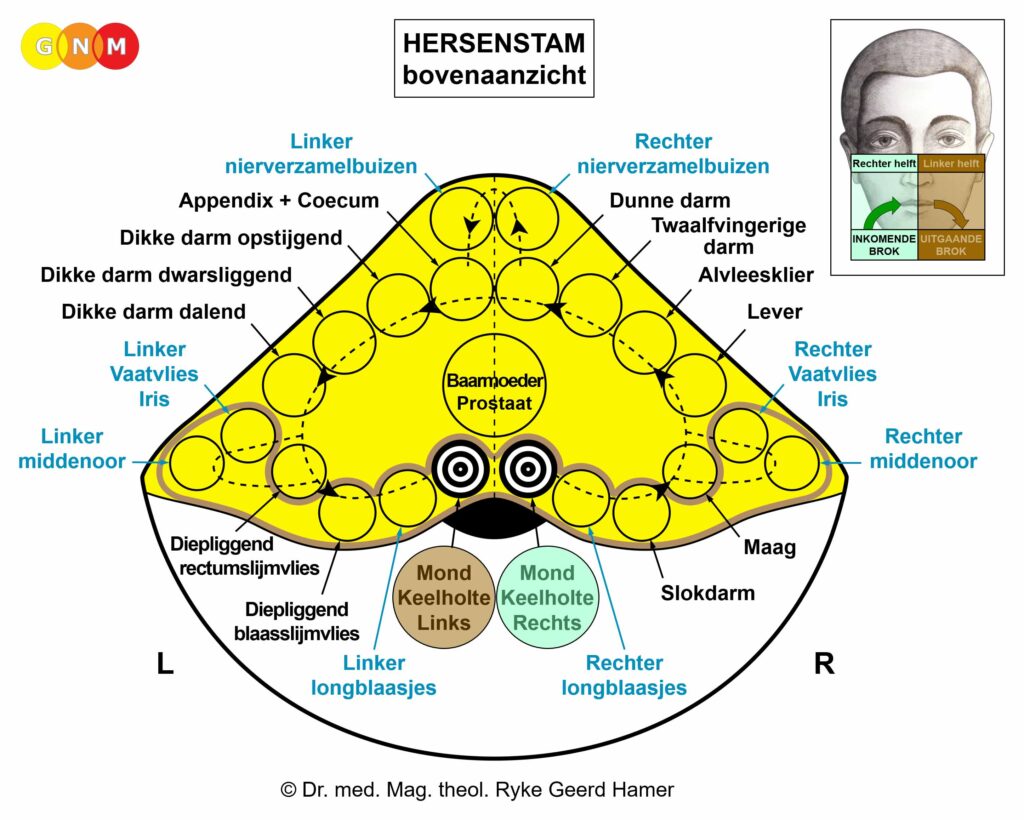
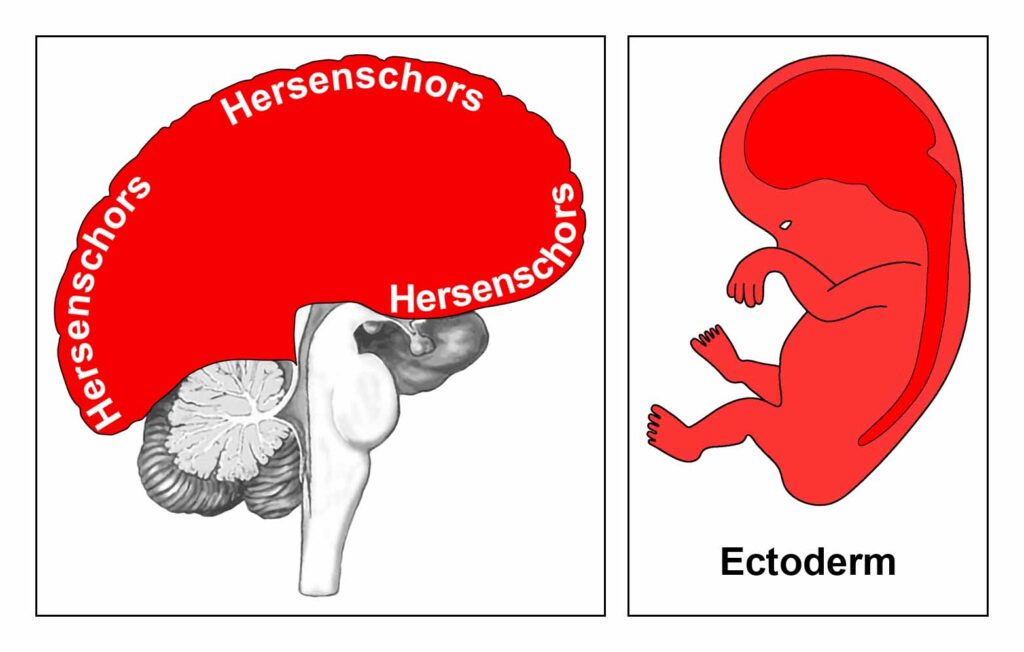
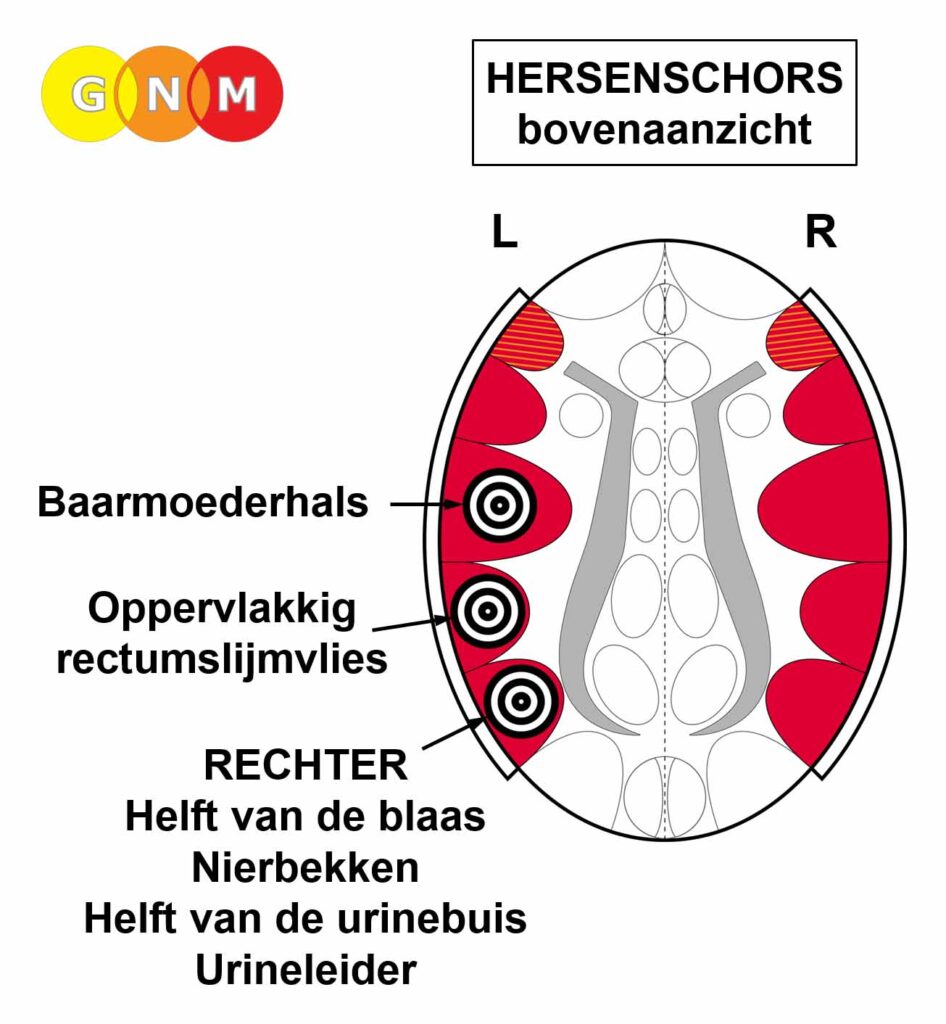
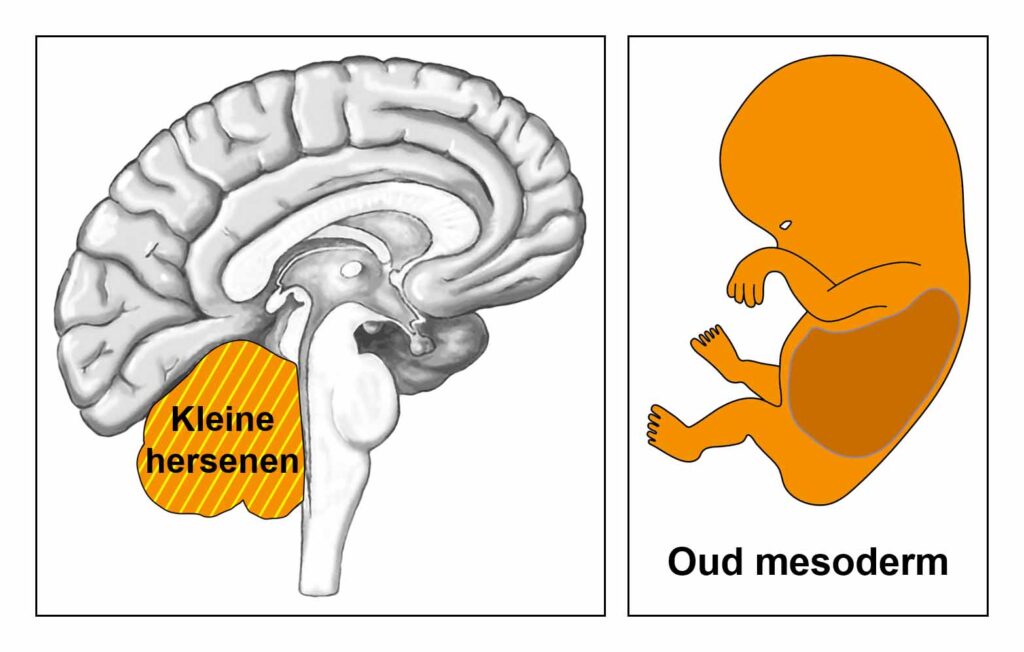
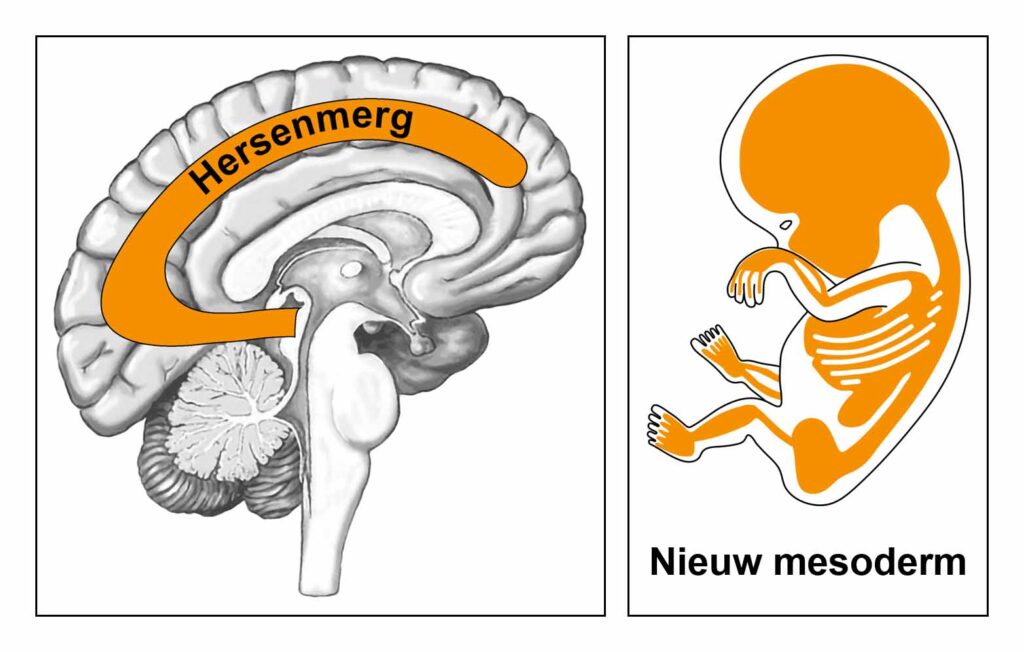
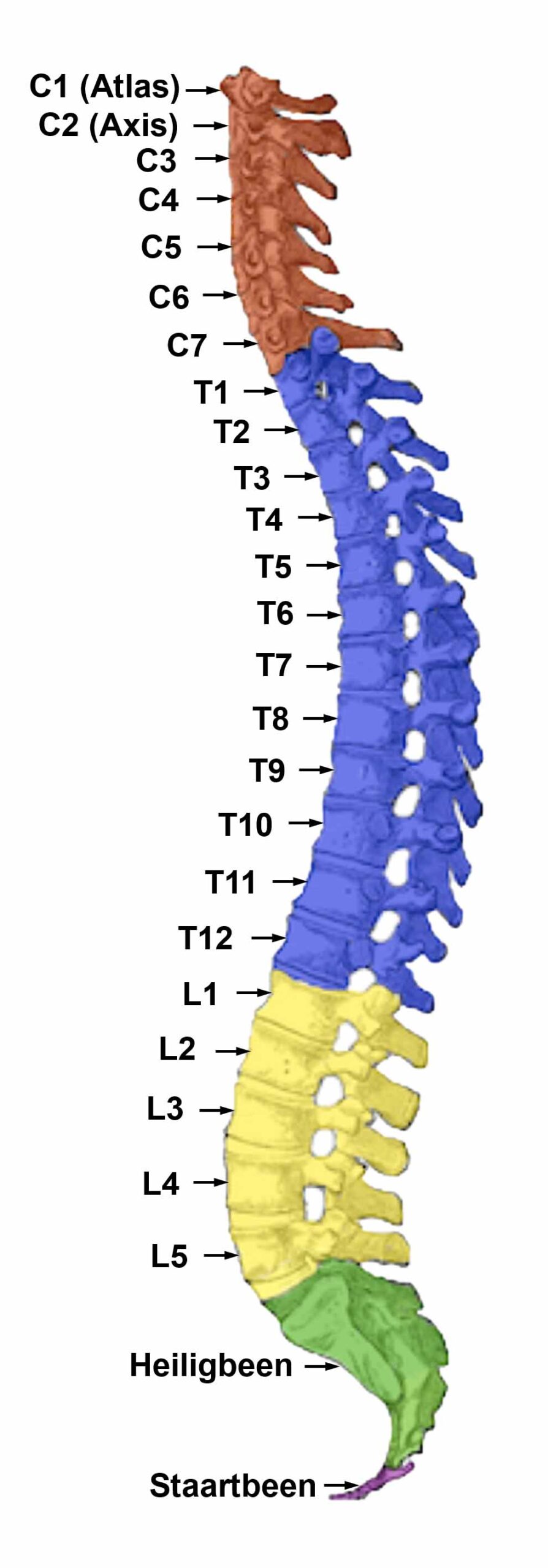
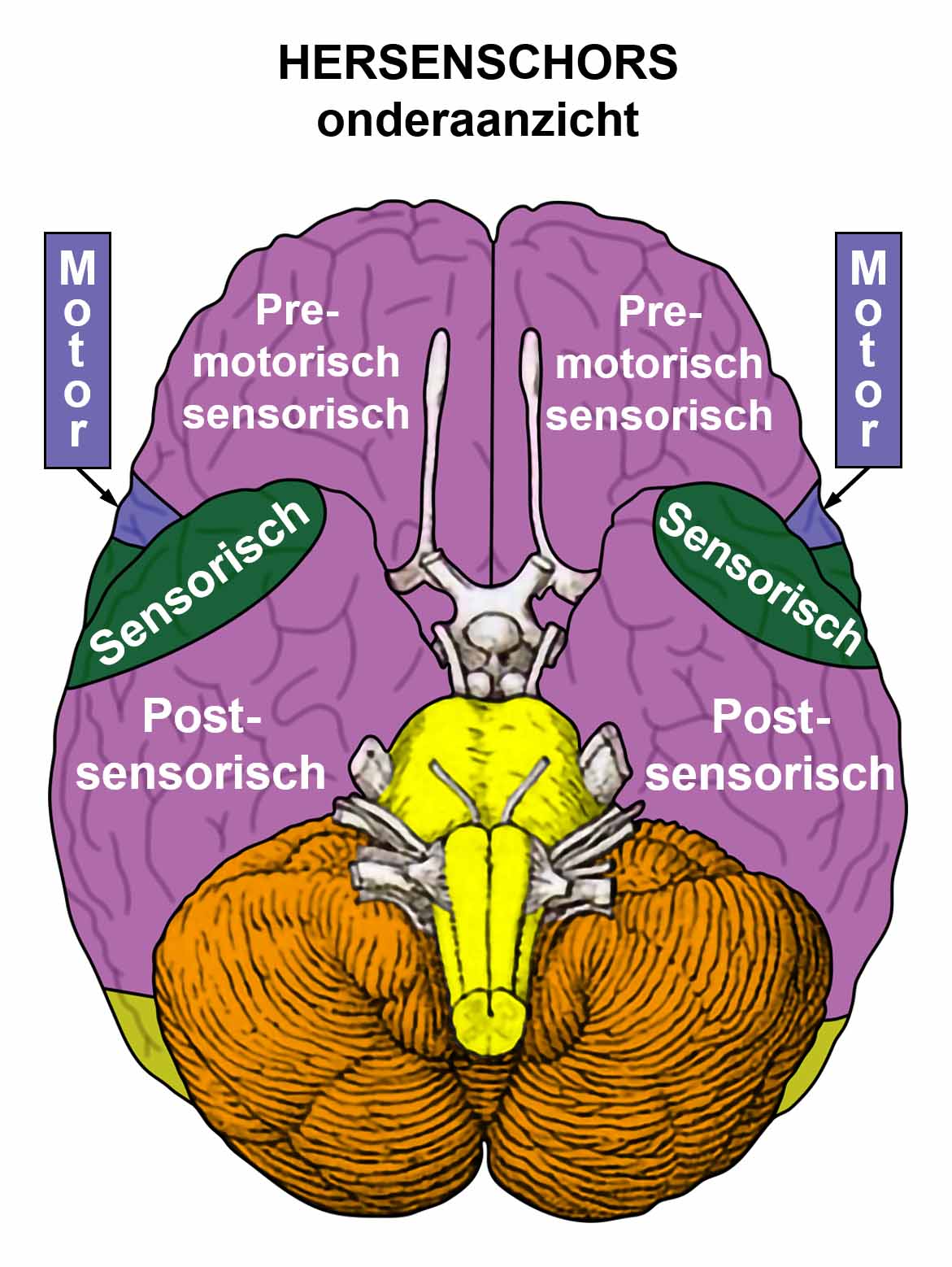
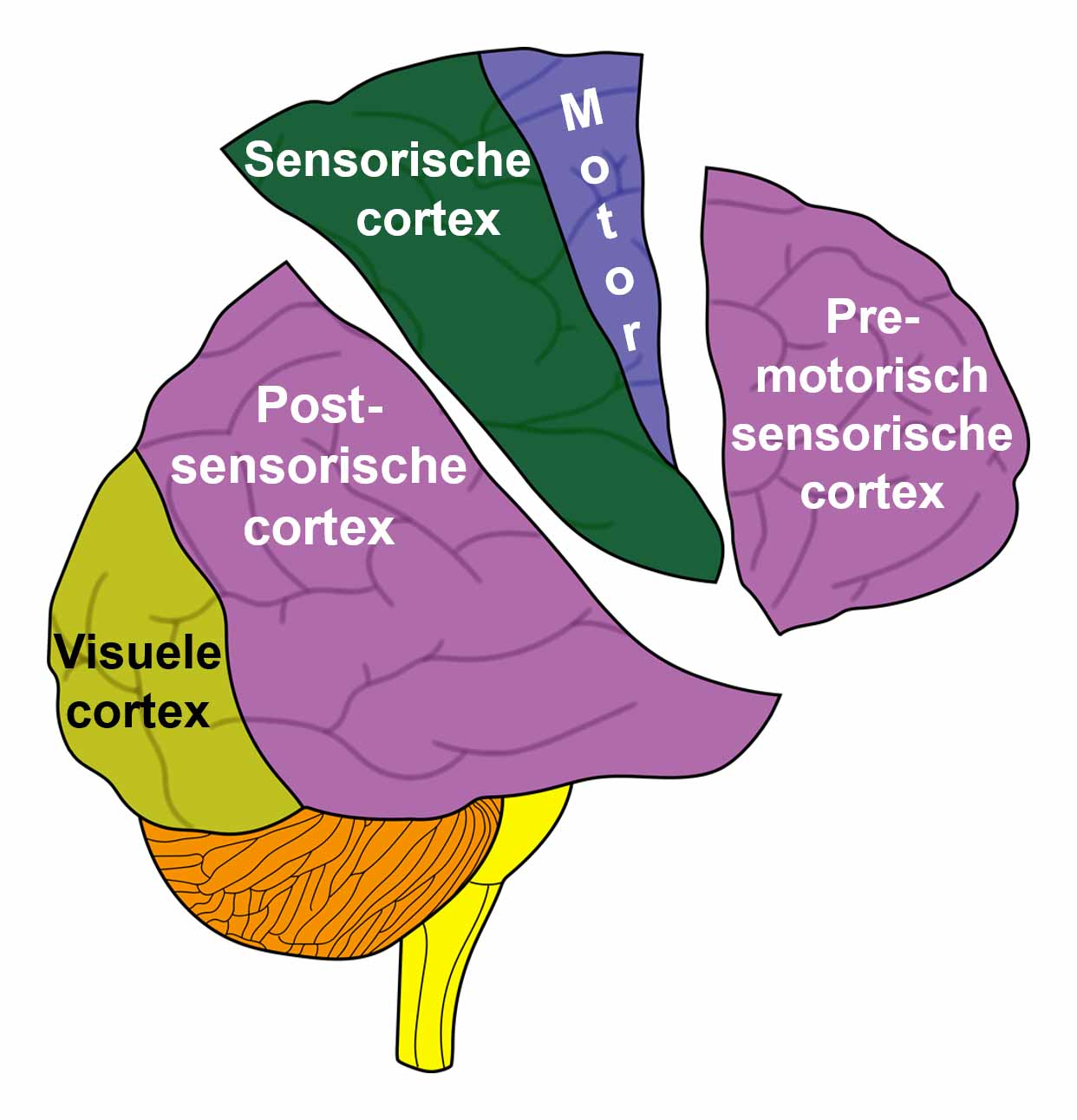
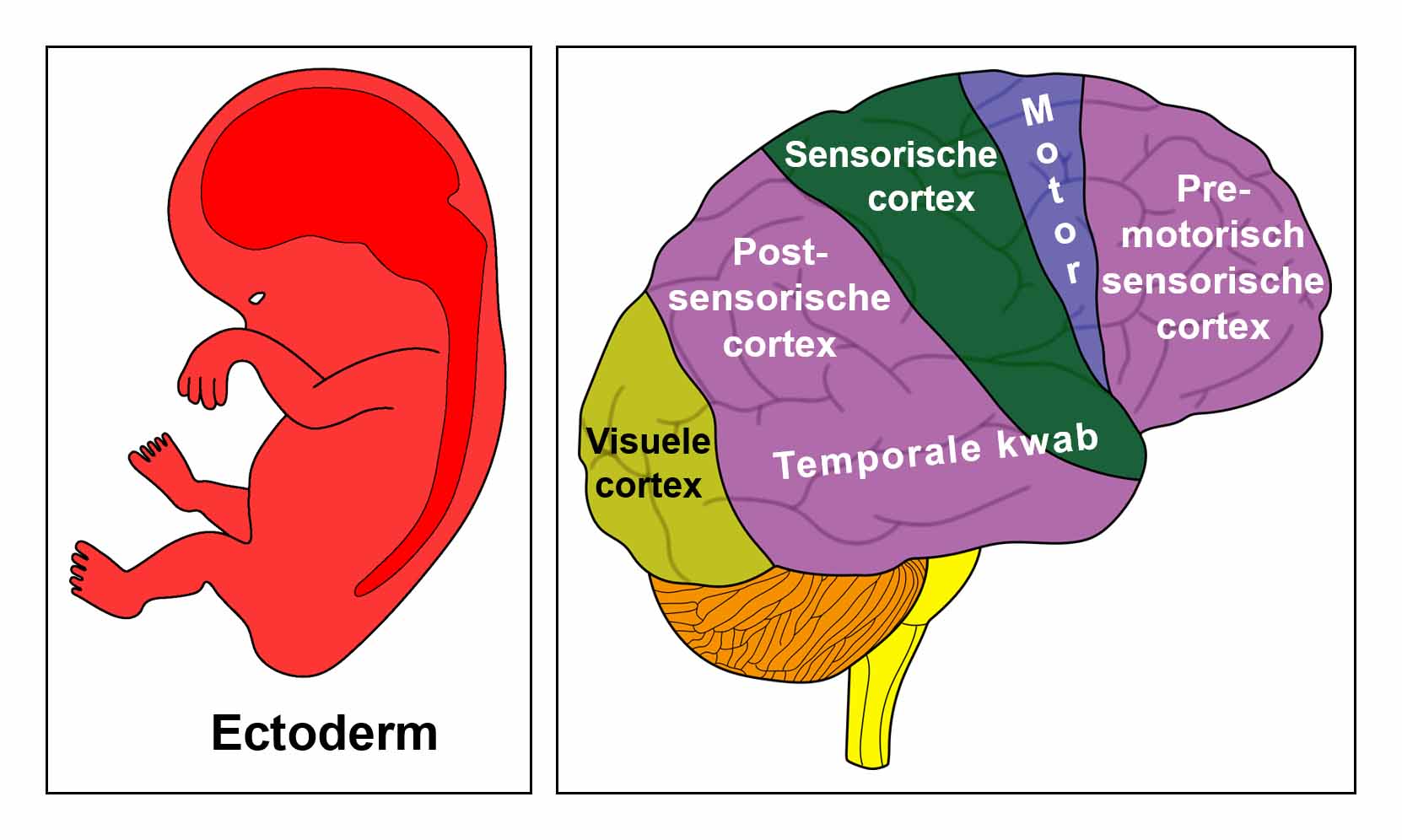
De gladde spieren van de darmen, de sigmoïd darm en rectum (bovenste deel), interne rectale sluitspier, nierbekken, urineleiders, blaas, urinebuizen, interne blaassluitspier, slokdarm, bronchiën, luchtpijp, strottenhoofd, baarmoeder, hartspier (atria), bloedvaten (incl. kransslagaderen, kransaderen, aorta, halsslagaderen en ondersleutelbeenslagaderen), lymfevaten, pupilspieren en ciliaire spieren zijn afkomstig van het ENDODERM.
De gladde spieren zijn onvrijwillige, niet-gestreepte spieren. Hun vermogen om samen te trekken veroorzaakt het verplaatsen van de voedselbrok (darmspieren), de bloedbrok (boezems, bloedvaten), de luchtbrok (strottenhoofdspieren, bronchiale spieren), de urinebrok (nierbekken, urineleiders, blaas, urinebuizen, interne blaassluitspier), de sperma-brok (prostaatgangen) en de lichtbrok (pupilspieren), door de specifieke organen met hun peristaltische beweging.
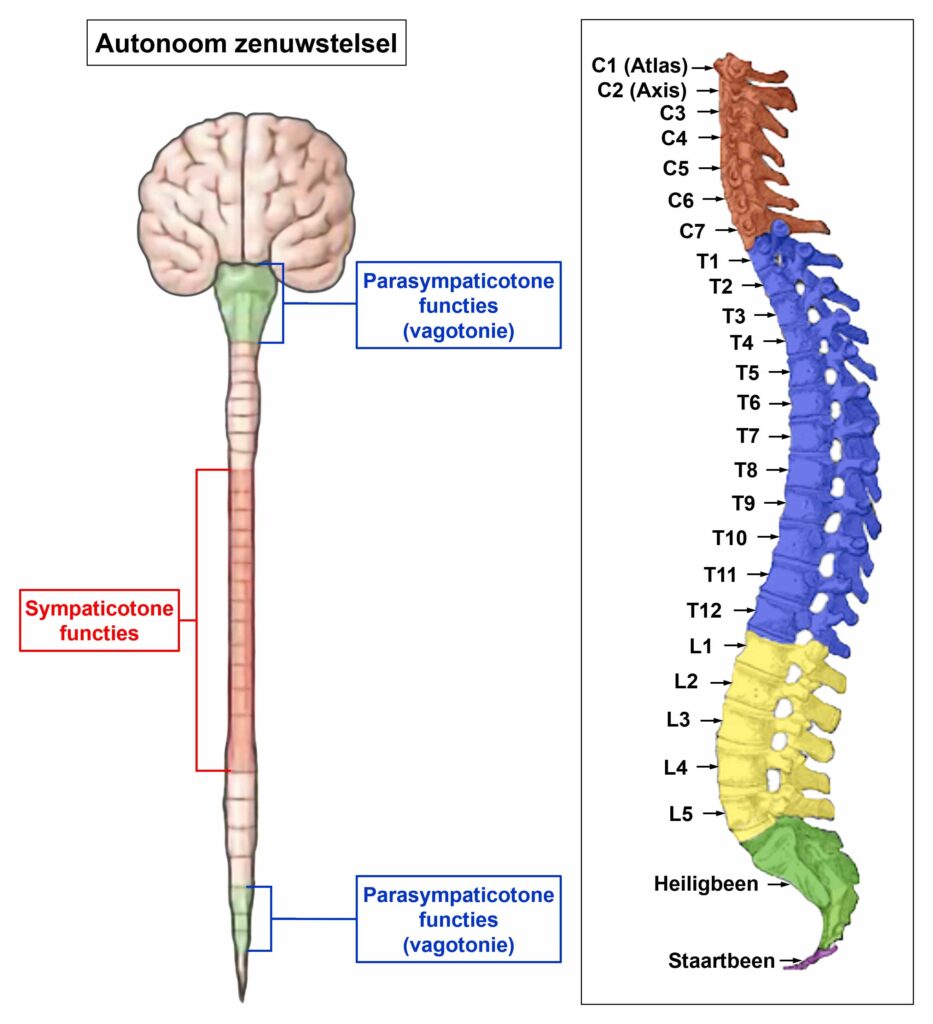
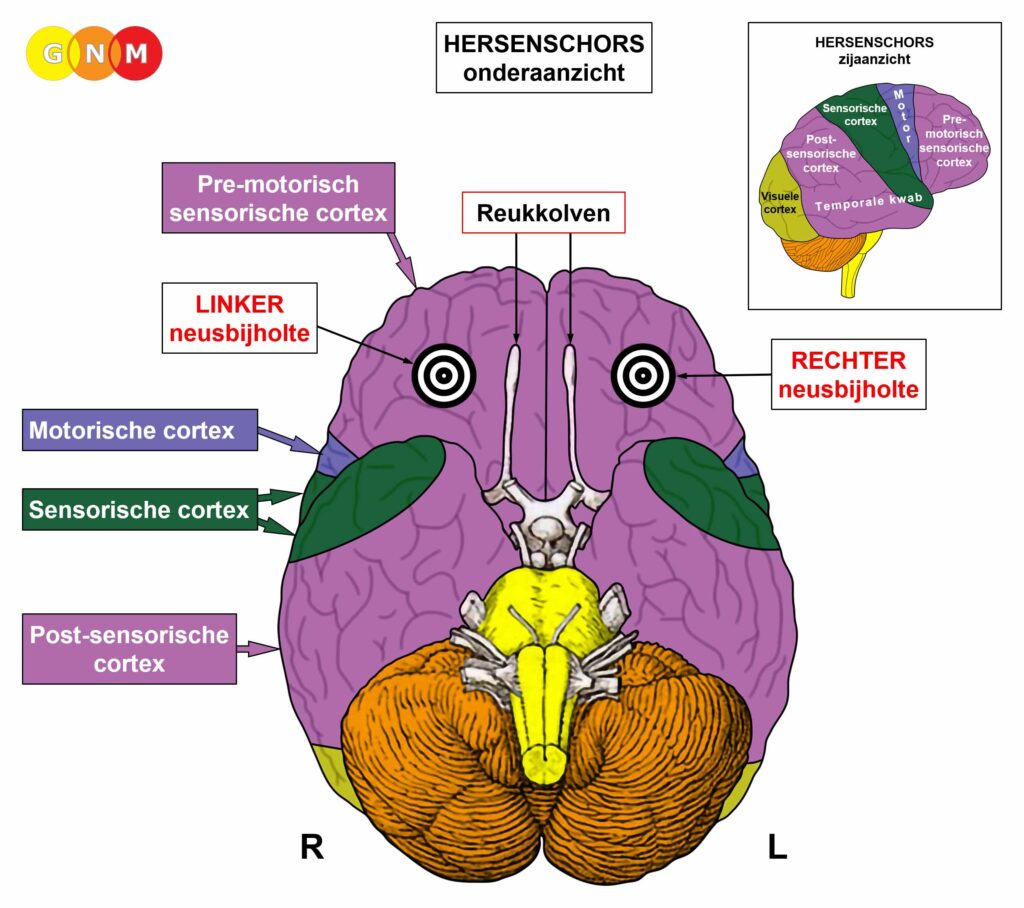
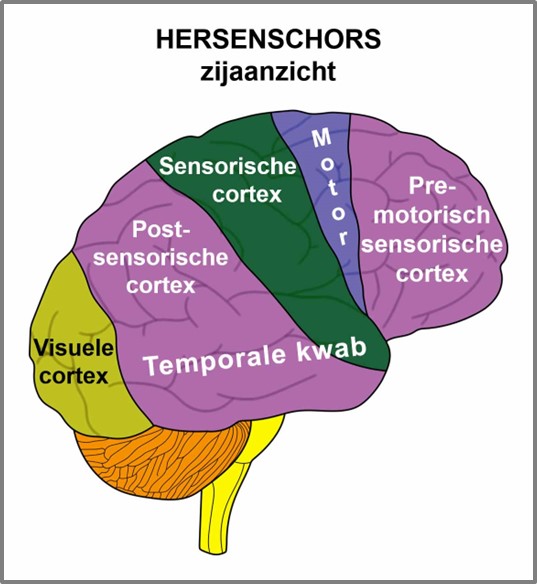
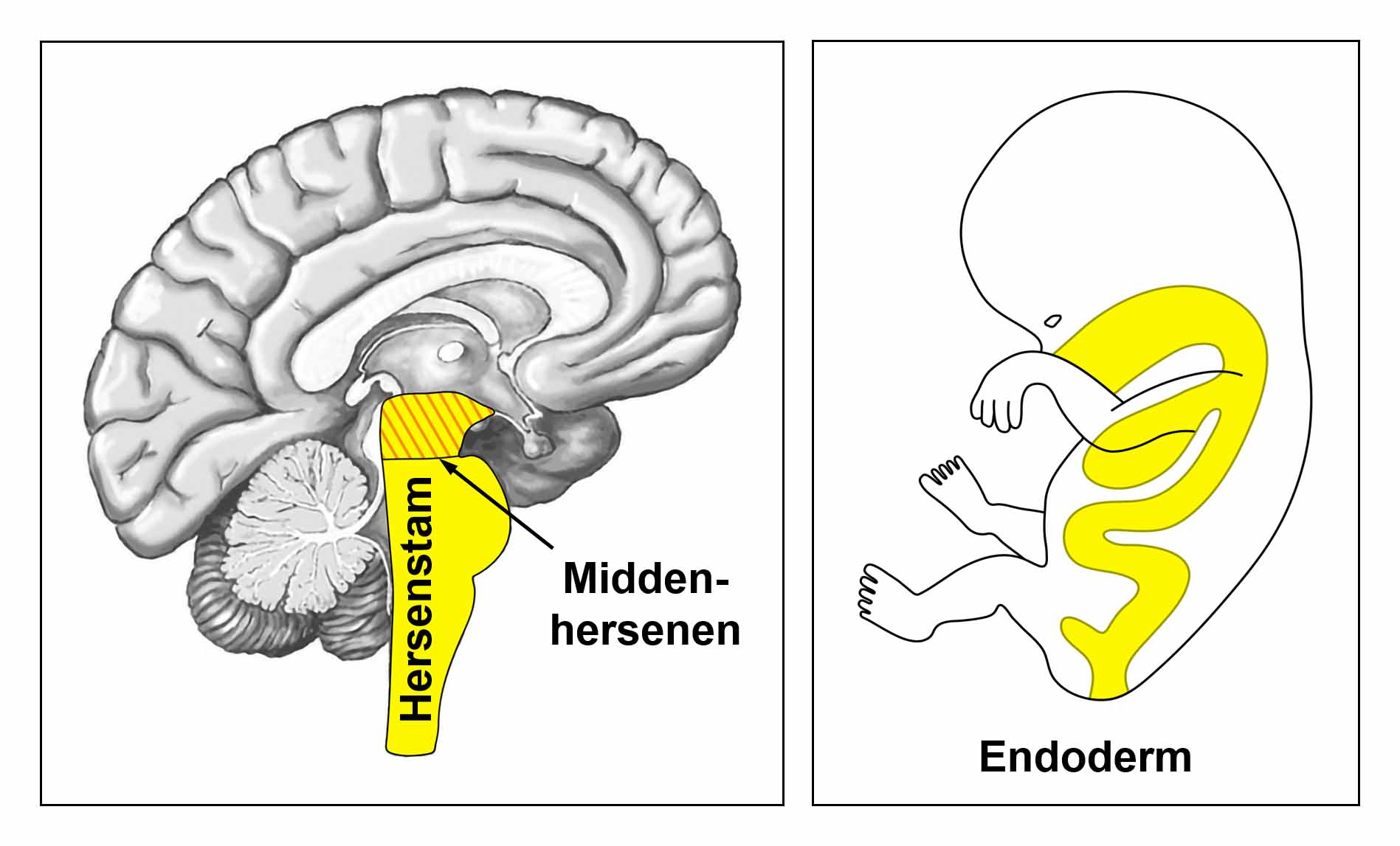
De gladde spieren worden aangestuurd vanuit de MIDDENHERSENEN, gelegen aan de bovenkant van de hersenstam. OPMERKING: De mannelijke en vrouwelijke kiemcellen worden ook aangestuurd vanuit de middenhersenen.
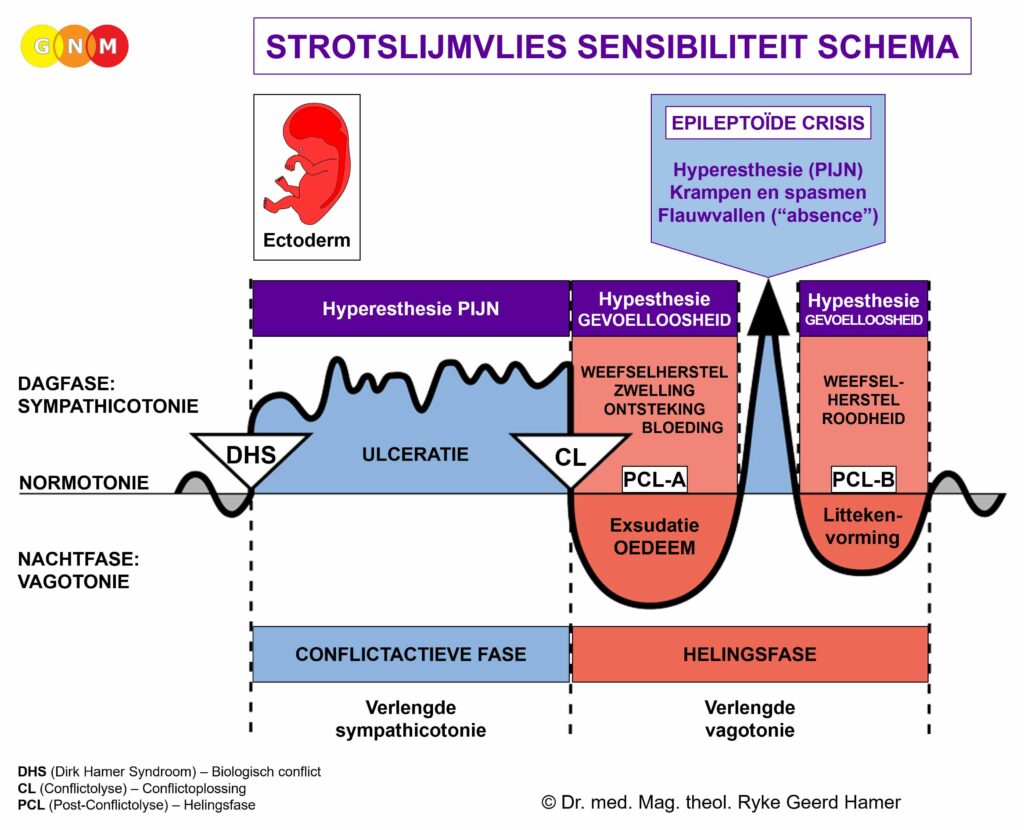
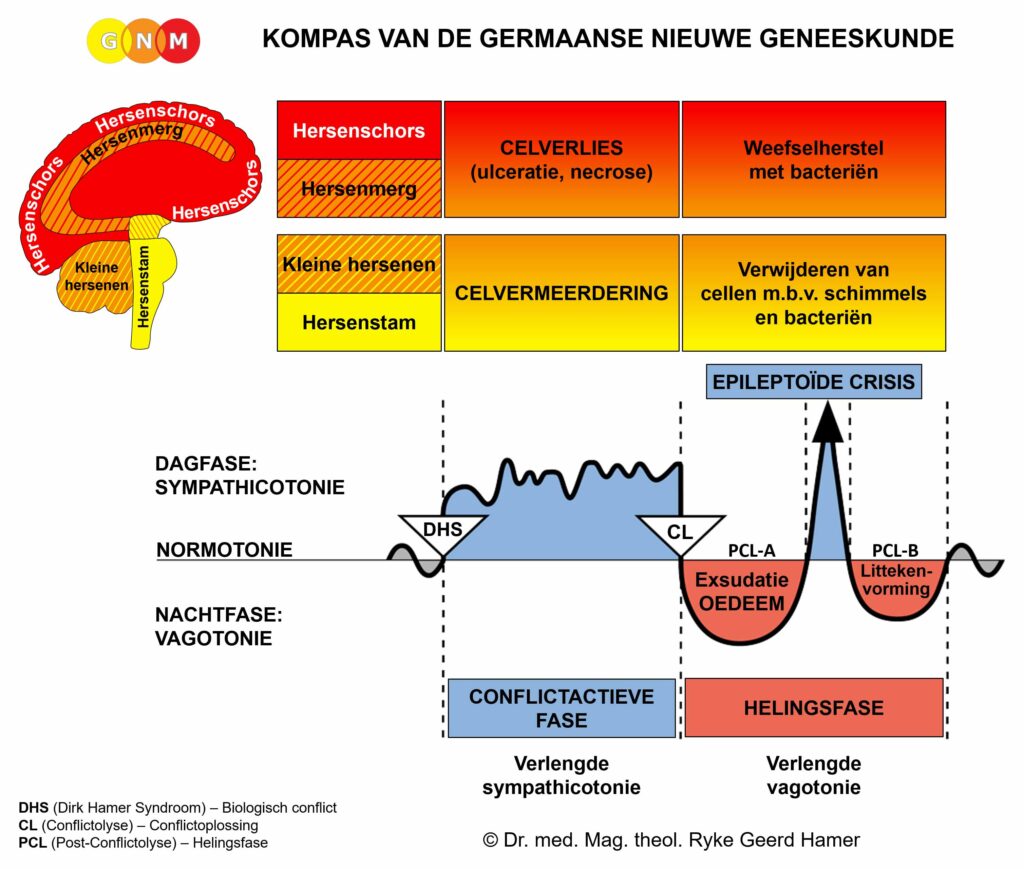
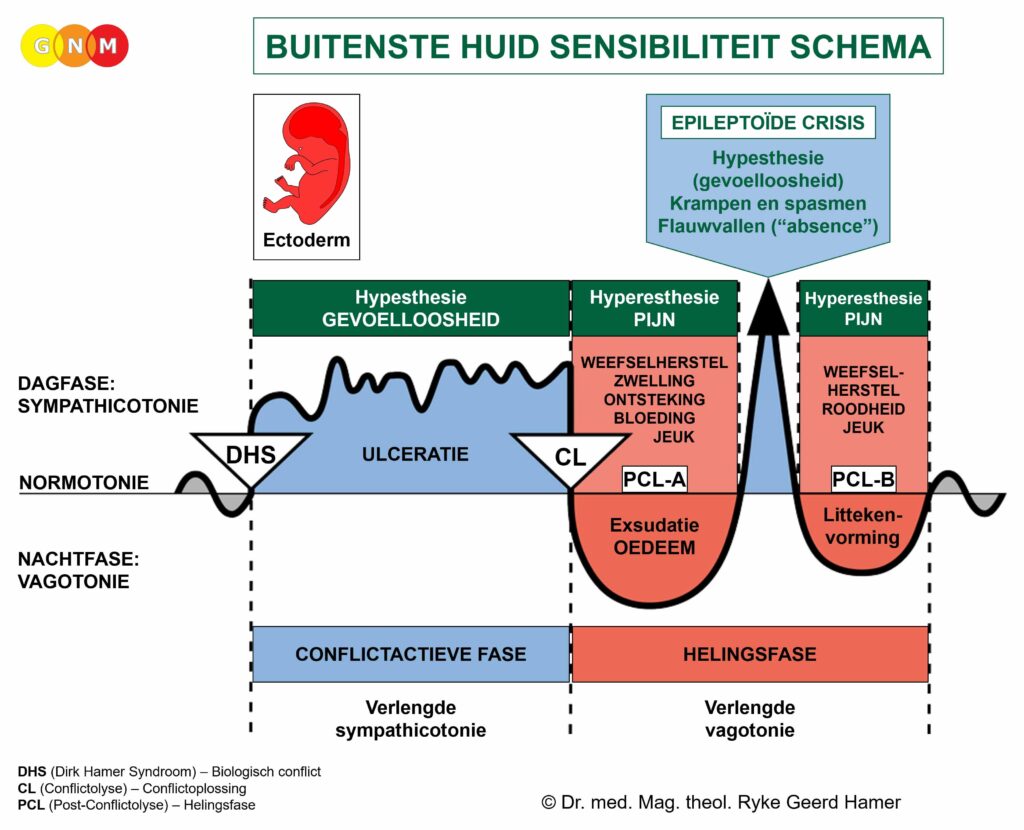
In het geval van een biologisch conflict genereren de gerelateerde spieren celvermeerdering tijdens de conflictactieve fase, met een toename van spiermassa en verhoogde lokale spierspanning (hypertonie) tot gevolg. In de helingsfase ontspannen de spieren. De Epileptoïde Crisis presenteert zich als spierspasmen. In de baarmoeder blijven de extra spiercellen aanwezig nadat de genezing voltooid is.